
The 15th of November 2022 was an unremarkable day that may have passed many people by, but this was the day that the world population reached 8 billion people. As many as 828M of those people will go to bed hungry tonight
Plant diseases caused by fungi and fungal-like organisms called oomycetes are serious threats to global food security and play a part in world hunger. These problems are also predicted to increase in a warming climate. Halting only the single most damaging of these diseases in the world’s top five food crops (rice, wheat, potato, maize, and soybean) would provide food for up to 4,2 million additional people per year globally, without the need for increased land use (Fisher et al 2012).
Current Policies within the EU
Healthy plants are essential, not only for our agriculture, horticulture, and other production systems, but also for our environment, landscapes, and biodiversity. Plant protection must be practiced without putting further strain on the environment, or human health for example, by avoiding hazardous synthetic pesticides where possible. This is the cornerstone of Integrated Pest Management (IPM) strategies, which the EU promotes through directives such as 2009/128/EC and the later amendment 2019/782. IPM focuses on pest prevention and alternative control methods that do not harm beneficial organisms such as pollinators, human practitioners or animals that depend on our plants. The EU Farm-to-Fork strategy highlights the need to transition to a fair, healthy, and environmentally friendly food system. The European Commission has also recently adopted a proposal for a new regulation on the Sustainable Use of Plant Protection Products. Under this regulation, the EC has set EU-wide targets for a reduction of synthetic chemical pesticide usage of 50% by 2030. However, for many crops, we still lack the scientific knowledge for the practical implementation of IPM, especially for plant diseases in open-field agriculture. Furthermore, there are limited biological control agents or other low-risk compounds that are registered as viable alternatives to synthetic compounds, especially for agricultural crops.
How can we bridge the gap between policy and practice for sustainable crop protection?
Scientific research for IPM needs to be strategically supported with long-term funding. We need to fund both excellent fundamental research and long-term applied studies to gain knowledge and deploy solutions for the control of plant diseases in our crops. We need to look to solutions through the lens of interdisciplinary and transdisciplinary research that is conducted in collaboration with stakeholders and that considers plant health in a holobiont context. EU projects such as Horizon 2020 EU-MSCA-ITN- 2017 PROTECTA (grant agreement number, 766048), and Horizon 2020 R&I program grant Organic plus (Grant Agreement No 774340) have helped us to bridge the gap between lab and field, and scientist and practitioner (Figure 1), but support is urgently needed for much longer projects, to allow time to evaluate plant protection strategies under field conditions including crop rotations with follow-up studies. Thus, we must be prepared to finance long term field trials over time periods covering 10-12 years to fully evaluate IPM strategies scientifically. However, in order to keep our crop plants healthy, we also need to better understand the fundamental biology of the interactions between our crop plants and their communities of both pathogenic and symbiotic microbes in our agroecosystems.

The plant as a holobiont
A new paradigm is emerging in plant health: the plant as a holobiont, or as a superorganism- a community of hosts and microbes (e.g., see Mannaa & Seo 2021, Figure 2). To take our scientific knowledge from the lab to the field and from policy to practice to sustainably control crop diseases, we need to connect our molecular plant- microbe interactions research to evolutionary biology and study these interactions at the ecological scale. We gain immense knowledge of the biology of plant disease and immunity by studying the molecular biology and biochemistry of the interactions between plants and their pathogens. We now know a very great deal about how the plant immune system works, and the discovery of resistance genes in plants has allowed and driven breeding for disease resistance in almost every crop grown today. Understanding the pathogenicity factors, or effectors that are produced by pathogens and how pathogens deploy such molecules to evade plant immunity is of fundamental importance to the development of crops with stable resistance to plant diseases. However, disease does not happen in a vacuum and neither does plant immunity. We now believe that the whole community of microbes, and other organisms, that are associated with plants can influence and support immunity or disease and therefore community approaches are needed to fully understand and enhance plant health sustainably in production systems.
Case study: IPM in the potato holobiont
Potato is the third most important food crop in the world in terms of production for human consumption. It is rich in vitamins and able to grow in a wide variety of climates, with lower water requirements than cereal crops. However, it is very susceptible to disease and 2M tonnes, or 7% of all the total usage, of fungicides, are used every year in the EU alone in potatoes. This is largely to control potato late blight, a disease caused by the aggressive and highly adaptable oomycete Phytophthora infestans and potato early blight, caused by the fungus Alternaria solani (Figure 3).
Late blight is the most economically important disease of potatoes and is efficiently controlled by synthetic fungicides. However, these often have to be reapplied frequently throughout the growing season and are an economic and environmental burden. Resistance breeding using wild relatives of domesticated potatoes has been ongoing since the 1950s, resulting in the widespread use of resistant cultivars in Europe. However, P. infestans can overcome this resistance within a few years. Furthermore, conventional potato breeding is a long and complex process, taking around 10 years before a new cultivar is commercially available.
It is clear then, that we need new tools in the IPM toolbox for potatoes and I believe that for IPM strategies that really work, we need to consider disease and immunity in a holobiont context (Figure 2) and importantly, we need much more knowledge on the biology of the microbes that cause these diseases and microbes that have potential as biocontrol agents against these diseases. For example, we have been studying the effects of both late blight and early blight on potatoes at the same time both in controlled conditions and in the field. P. infestans is a hemibiotrophic pathogen and as such needs living tissue for early infection. It therefore utilises a “stealth attack” mode of infection, where it deploys an array of effector molecules to subvert plant immunity and reprogram plant cells for disease. A. solani on the other hand is a necrotrophic pathogen, operating a “brute force” attack strategy whereby plant cells are rapidly killed, and the resulting nutrients are consumed by the pathogen. We know a lot about the biology and molecular weaponry used by these two pathogens from detailed studies of each of them individually. However, in agricultural fields, we have seen that potatoes may face both these pathogens simultaneously. Interestingly, our studies indicate that the presence of P. infestans appears to help A. solani and more severe disease symptoms occur when both pathogens are present together, or where P. infestans arrives first. On the other hand, when A. solani arrives first, it is very difficult for P. infestans to later attack since the potato immune system has already been activated and tissue has begun to die (Brouwer et al 2022). We are currently investigating the molecular determinants of these tripartite interactions, to see how potato responds to these two pathogens together at the molecular and biochemical level and which effectors or other factors are deployed by these pathogens when they are competing or collaborating to cause disease in the same host plants.

We have also identified wild Solanum species in Sweden that can act as overwintering hosts for the late blight pathogen. Isolates of P. infestans that have overwintered in the rhizosphere of Solanum dulcamara are more pathogenic on potatoes than isolates that have previously only infected potatoes (Vetukuri et al 2020).
Given the propensity for P. infestans to quickly adapt to new potato cultivars and the economic and environmental burdens of repeated fungicide usage, as well as the build-up of fungicide resistance in A. solani populations, effective biocontrol solutions would be highly beneficial in potatoes. However, within the EU, only three biocontrol products are licensed for use against late blight in potatoes. These are Taegro, a formulation of the bacterium Bacillus subtilis var. amyloliquefaciens, Polygandron, a formulation of the oomycete Pythium oligandrum and Sonata, a formulation of the bacterium Bacillus pumilus. Under laboratory conditions, we have observed that the mycoparasitic oomycete Pythium oligandrum can effectively parasitise the late blight pathogen within 48-72 hours and it secretes an arsenal of destructive enzymes and effectors to do so (Horner et al 2012; Liang et al 2020). Under greenhouse conditions, P. oligandrum is also able to significantly reduce early blight disease on potatoes (Stridh et al 2022), however, the effects diminish in field trials. In small-scale field trials treated with hand spraying disease control effects are still observed with several biocontrol agents, but in large-scale trials treated with tractor sprayers, the effects are almost completely lost (Stridh et al 2022). These experiments suggest that factors such as environmental conditions including UV radiation and rainfall may hamper the effectiveness of biocontrol agents (BCAs) under field conditions. The decrease in effectiveness from small-scale to large-scale field trials, where the application technology was changed from hand spraying to tractor spraying suggests that application technologies need to be adjusted for BCAs in open-field agriculture. Current practice is to utilize existing pesticide spraying technologies however, these may not adequately distribute BCAs or allow them sufficient contact with the pathogens themselves.
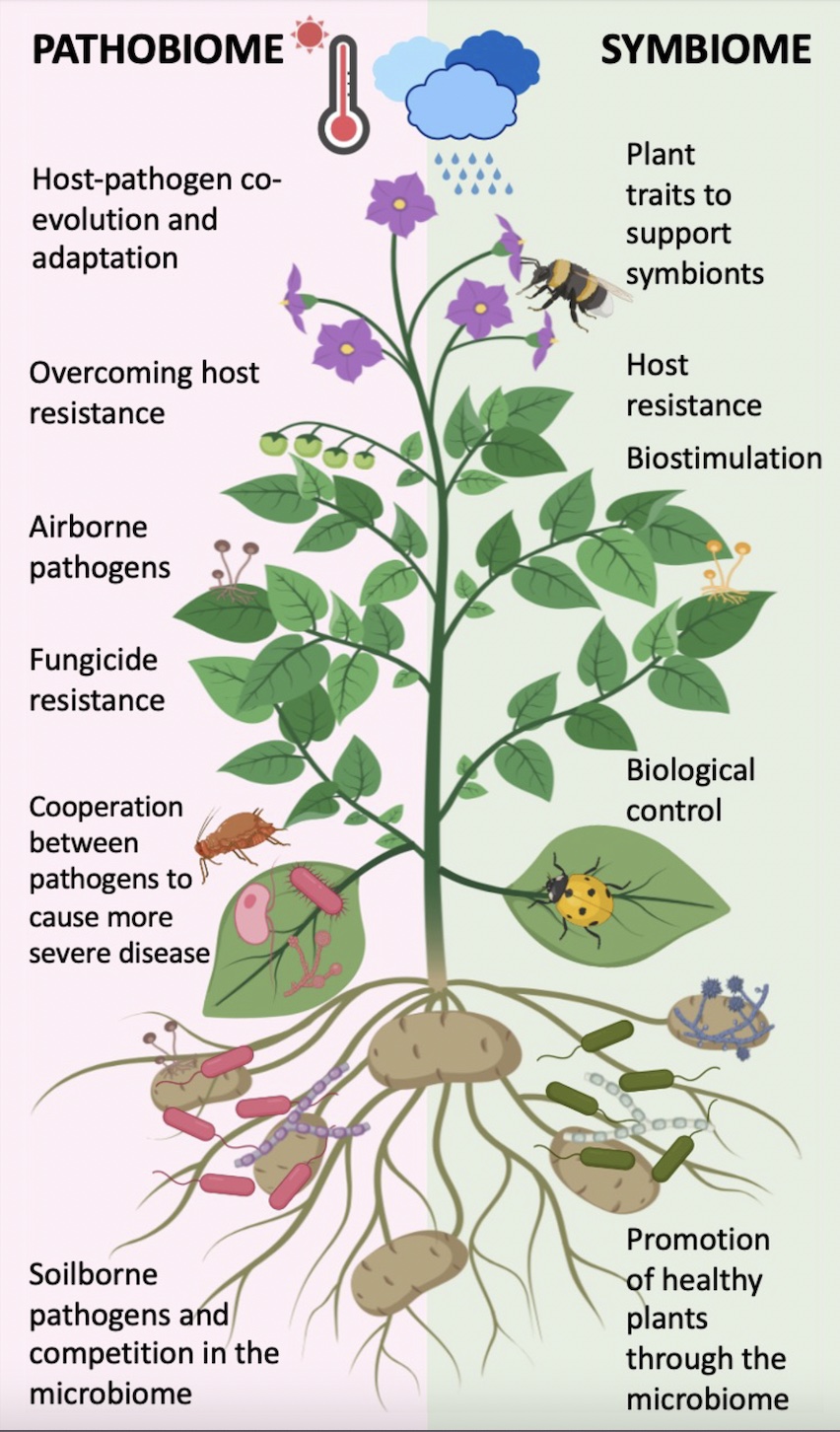
Pathogens or organisms with negative effects on potato health are in pink and those with positive effects in green.
As well as biological control, where the pathogen is directly or indirectly targeted by a living BCA, other promising alternatives to synthetic pesticides include the use of plant resistance inducers or PRIs. These are agents that lead to improved plant protection by inducing the plant’s own defense mechanisms and are often biologically derived. For example, β-aminobutyric acid (BABA) has been shown to induce resistance against P. infestans in potato cultivars and their wild Solanum relatives (Lankinen et al 2016). However, in a similar manner to BCAs, more research is required to test the best application technologies to deliver these in open-field agriculture and to assess their effectiveness under different environmental conditions.
For effective use of both BCAs and PRIs further investigations into their interactions with host plants at the genetic level are needed. For example, we have been breeding potatoes for resistance to late blight and for other desirable traits such as yield for decades, however, we have neglected to select plant genotypes that are inducible by PRIs or that are best able to host symbiotic microbes that act as BCAs. A recent study in sugar beet identified host genotypic differences in biostimulation by the symbiotic microbe Trichoderma afroharzianum T22 (Schmidt et al 2020) and several studies suggest responsiveness to PRIs is plant genotype dependent (reviewed in Sandroni et al 2020), although it may also be influenced by the genotype of the pathogen too, which will be challenging to address in future breeding programs.
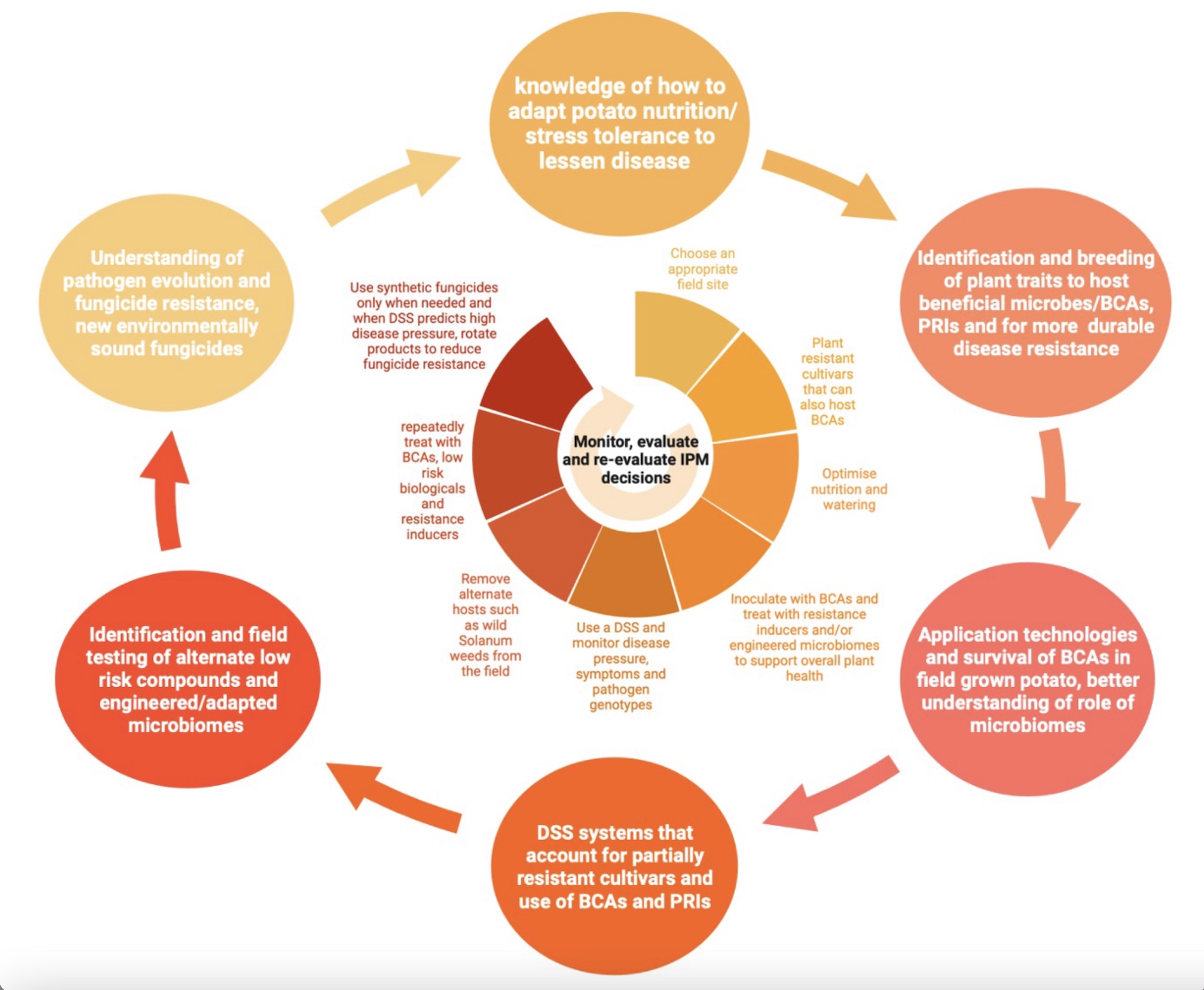
Scientific knowledge gaps to be addressed (outer circles) to support implementation of holistic IPM approaches in potato (inner circle)
Along with hosting specific beneficial microbes, the plant as a holobiont is also a community of microbes or microbiomes that work in concert with the host plant itself to support plant health. The richest of these microbial communities are the rhizosphere and root microbiomes, which have been compared to the human gut for their roles in acquiring nutrients, removing waste, and supporting overall plant health (Ramirez-Puebla et al 2013). Our understanding of plant microbiomes is developing at a rapid pace, largely due to the development of affordable and accurate large-scale sequencing technologies in recent years. We are beginning to understand that plants shape their own microbiomes and will take on microbiota for different needs at different times. For example, one recent study showed that the model plant Arabidopsis thaliana will recruit the fungus Colletotrichum tofieldiae when it is short of soluble phosphate and wants help. When phosphate is abundant it repels the fungus. In most other situations this fungus is a devastating pathogen (Hiruma et al 2016). However, how plant microbiomes are recruited, structured, or modulated by environmental factors is not yet well understood. In potatoes, studies have shown that soil and cultivar type both influence the bacterial microbiome (Inceoglu et al 2011) and that fungal and bacterial microbial community structures change during different growth stages of potatoes (Hou et al 2020) and that these may affect or be affected by inoculation with BCAs (Roquigny et al 2018; Song et al 2021).
The way forward for sustainable IPM cultivation
There are still a lot of knowledge gaps for the development of effective and sustainable IPM in open field cultivation of crops like potatoes, as summarized in Figure 4, (outer circles). However, if we can breed potatoes to better host beneficial microbes including BCAs such as P. oligandrum or Trichoderma species, at the same time as breeding for both stimulation of plant immunity through the application of PRIs and durable disease resistance, e.g., through stacking of multiple disease resistance genes or gene editing of susceptibility genes we will have a firm foundation for the prevention of diseases such as late blight and early blight in potato. Since these pathogens are highly adaptable and the selection pressure in monoculture fields of genetically identical potato clones provides the impetus for rapid evolution in these pathogens, we also need to be ready with new tools for the treatment of these diseases. Thus, more research to identify how best to deploy BCAs to withstand the harsh environment found in open field systems, and to deliver both BCAs and PRIs to the right place in the plant to allow contact with the pathogens should support better biological control of these diseases. The next steps will be to integrate BCAs and PRIs into decision support systems (DSS) for farmers, modelling the best time to apply these control measures to the potato crop. A better understanding of pathogen evolution for fungicide resistance and host-pathogen co-evolution for disease or immunity is also required to develop control strategies that will be durable in the field without an over-reliance on synthetic pesticides, as shown in the inner circle of Figure 3, which depicts what potato IPM could look like in the near future. The combination of these measures should make it possible for us to cut down our reliance on synthetic pesticides in crops like potato and more fully embrace IPM in open field agriculture.
Read and download the full eBook and list of references here ‘Towards sustainable control of plant diseases’